
The starting point for this discussion is the development of vertebrate fish nearly 500 million years ago. Fish swim; they do so by curving their spines alternately left and right, like this:
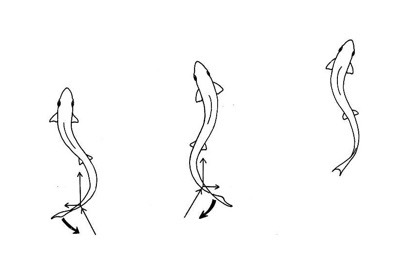
When the first fish/amphibians crawled up onto the land, they used the same basic technique to crawl: they curved their spines to the left, placed their right front foot down, then curved their spines to the right, placing their left front foot forward, like so:

However, crawling is slow and inefficient, because you waste lots of energy dragging your tummy along the ground. The next step was to move the legs from the side to underneath the body; land animals graduated from crawling to walking. This greatly improved both the speed and energy efficiency of walking.


At first, the legs were merely moved outwards and downwards somewhat, as with the lizard on the right of this picture. Later on, the legs moved completely underneath the animal and gained length to further increase speed and efficiency:

Gaits
But an animal with four feet can do more than simply walk; it can change the order and speed with which it moves its legs. These different schemes for moving one’s legs around are called gaits, and there are lots of possible combinations of motion of those legs, so there are lots of different gaits. However, there are only a few basic types of gait, as shown with horses:

There’s a big problem with equipping an animal with multiple gaits: how can all the neurons required to control the muscles be wired up? That wiring will have to be very complicated. It’s easy enough to come up with the connections for crawling. Those connections can be programmed right into the DNA so that the creature can crawl immediately after birth:

But equipping an animal with, say, four different gaits poses a much more complex problem. Each neuron has to be wired up so that, depending upon the gait that the creature has chosen, it fires at different times relative to the other neurons. This requires the addition of extra timing neurons to delay the firing of different neurons depending upon the gait. This can be very complicated — too complicated to be hardwired into the genes.
It gets worse: walking is the slowest of the gaits. Other gaits, such as running, are faster and so require more precise timing of the neurons. For example, watch this video of a cheetah running (the relevant part is from 0:57 to 1:23).
The cheetah’s front feet hit the ground just 10 milliseconds apart. I think it’s obvious that, if the cheetah’s muscle timing was off by as little as one millisecond, those feet would be misplaced and the cheetah would stumble. In other words, the cheetah’s overall timing of nerve cells must be accurate to better than one millisecond. This presents us with a new problem: the average neuron has a firing precision of only about 11 milliseconds. The cheetah needs better than 1 millisecond precision yet its neurons are only good to 11 milliseconds. There is a way around that problem: bunch together lots of neurons and average their signals together; the more neurons you add, the more precise will be their average firing time. To understand why that is, imagine a little experiment: you want to time with great precision the exact moment when a runner crosses the finish line, and you don’t have any electronic equipment, just a lot of stopwatches. You could have one observer who measures with the stopwatch the instant that he sees the runner crossing the finish line, but that observer might be early or late. Suppose instead that you had a hundred observers all crowded together at the finish line, and each of them pressed their stopwatch button the instant they saw the runner cross the finish line. Some of the observers would be a shade early and some would be a shade late, but if you take the average of all their timings, you’d get something that is more accurate than what you’d get out of a single observer. Substitute neurons being fired for people pressing stopwatch buttons and you get the idea.
So not only must we come up with a way to trigger muscles in different timings and ordering depending upon the gait, but we also need a vast number of neurons doing all this to get the timing precise enough for fast running. In other words, we’re going to require a vast and complicated wiring diagram for the nervous system of any animal that wants to use multiple gaits. The obvious way to give that wiring diagram to the animal is to encode it in the animal’s genes. That solution runs into a brick wall: the amount of DNA required to encode all that information is prohibitively high. You’d need lots of extra chromosomes to handle that kind of system. Worse, it would take a long time for natural selection to get all the wiring genes tuned up properly. And one little mistake in one gene would reduce a sleek cheetah to a shambling clod.
But then, sometime around 150 million years ago (in my opinion), something remarkable happened. A new system, based on three components, appeared. I don’t know how it evolved, or what its antecedents were, but I believe that the three key components came together roughly 150 million years ago.
The first component was the development of a new and improved neuron. This neuron could transmit signals and respond to large numbers of inputs, just like its ancestor neurons. But this new neuron had one extra capability: it could alter its response to inputs based on what happened to it. It could change the weights that it assigned to different inputs, putting more emphasis on some inputs and ignoring others. In other words, it could change its wiring after the creature was born.
The second component was the development of a new behavior based on random trial and error. Animals would wiggle their limbs around in all sorts of crazy ways. This new behavior enabled animals to try out all sorts of activities, most of which were spastic, but a few of which might prove useful.
The third component was a new system for wiring together neurons into a nervous system. Instead of having every connection planned in advance, the DNA simply programmed neurons to connect to just about everything they could reach. In other words, animals were born (or hatched) with an overconnected nervous system.
We have names for the first two components: learning and play. By playing at moving around, animals were able to teach themselves how to run, walk, gallop, trot, and so on. Learning and play were invented simultaneously, as two parts of the same development. It’s ironic, isn’t it, that nowadays we wonder whether play can be used to improve learning when in fact, play and learning are really two sides of the same evolutionary coin.
Aside: this view of learning and play isn’t really that unorthodox: it’s just a neurological analog of what genes do in natural selection. Accidental mutations generate variations in the genes, and those variations are tested against the demands of the environment, rejecting the mutations that don’t help, and preferring the mutations that do help. If you want to anthropormophize the process, think of Mother Nature “playing” with the genes for a species, trying out different variations. Most of those variations produce stupid, deformed, slow, or otherwise deficient creatures. But every now and then Mother Nature hits the jackpot with a new and better set of genes.
In the neurological analog of this process, play generates the variations in locomotor behavior, which are then tested, not against the environment per se, but against the animal’s expectations of efficient and speedy movement. Play is the random generator used to refine the connections in the nervous system. The two primary differences between the two systems are 1) that the genetic evolutionary process requires many generations to function, while the neuronal evolutionary process -- again, what we call learning -- takes place over a time scale of days to months; and 2) that the evaluation of effectiveness is external (the environment) in the case of genes, but internal in the case of neurons -- there’s an internal evaluation process that decides whether the creature is moving well.
I therefore maintain that learning and play developed to take fullest advantage of the opportunities offered by multiple gaits. Once the basic technology of learning and play was in place, it was applied to many other areas of mammalian behavior. This process, of course, required millions of years to develop.
Carnivorous mammals developed an especially complex system of play: